Andrey A. Gurtovenko1, 2
1Microplastics Research Center, Yaroslav-the-Wise Novgorod State University, Veliky Novgorod, 173003, Russia
2Department of Physics, St. Petersburg State University, St. Petersburg, 198504, Russia, E-mail: a. gurtovenko@gmail. com; Web: biosimu. org
ABSTRACT
Microplastics, tiny fragments resulting from the degradation of plastic waste, are abundant in water, air, and soil and are currently recognized as a global environmental problem. There is also growing evidence that nanosized microplastics can be hazardous to living species. Unlike most experimental methods, computer modeling is particularly well suited to studying the effects of such nanosized microplastics. Here we use atomic-scale computer simulations to explore for the first time the impact of polymer nanoparticles on model cell membranes containing cholesterol, an essential component of membranes of eukaryotic cells. Our findings clearly show that, once a polymer nanoparticle (~4. 0 – 4. 5 nm in diameter) partitions into the membrane interior, it alters both the lateral and transverse distributions of cholesterol. Polymer chains induce the formation of domains depleted in cholesterol and also promote cholesterol flip-flops. These changes could have severe consequences for living cells as they affect the fluidity and the phase behavior of membranes, the transmembrane lipid distribution, and the ability of membranes to balance the stress.
INTRODUCTION
The term microplastics was introduced 20 years ago to describe tiny fragments resulting from the degradation of plastic waste abundant in the oceans. 1 At present, microplastics are widely recognized as an important contaminant of water, air, and soil and are considered a global environmental problem. 2 There is growing evidence that microplastics can be hazardous to living species, with size being a major factor for such a threat. 3, 4 According to the definition, microplastics represent polymer particles that are less than 5 mm in diameter. However, microplastics-induced cellular toxicity is generally expected for nanosized plastic particles (so-called nanoplastics) because of their ability to translocate through cell membranes. 5 For instance, microplastics with a size of 500 nm and smaller can enter cells via endocytosis pathways. 6 When the size of microplastics approaches the thickness of cell membranes (5-10 nm), a plastic particle can cross the membrane directly via passive diffusion. 5 Importantly, the smaller the microplastics, the more challenging their detection and characterization due to resolution limitations of experimental techniques. On the contrary, as far as computer modeling is concerned, the smaller the better. Therefore, computer modeling could serve as a valuable source of information regarding the impact of nanosized microplastics on cells and cell membranes.
First computational studies on microplastics-membrane interactions employed simplified coarse-grained models. This approach allowed one to overcome the limitations of time and length scales, which are inherent to computer modeling of complex molecular systems. 7-9 In the landmark study by Rossi et al, 7 the authors used coarse-grained computer simulations to follow the permeation of polystyrene nanoparticles into model lipid membranes and their subsequent dissolution in the membrane’s hydrophobic core. They found that a polystyrene nanoparticle entered the model membrane and perturbed its structural and dynamical properties upon dissolution. 7 A more accurate description requires models of higher resolution, which are computationally much more expensive. Nevertheless, recently the corresponding atomistic computer simulations started to emerge. 10-14 While these studies provided a detailed, atomic-scale insight into the interactions of microplastics with model cell membranes, the focus by far was on simple, single- component phospholipid bilayers. However, the composition of the lipid matrix of plasma membranes is far from that simple. 15 Besides phospholipids, cholesterol is one of the most abundant components of cell membranes; the cholesterol concentration typically amounts to 20-30 mol % but can be as large as 50 mol %. 16 Relatively small, stiff molecules of cholesterol are essential for the properties of biological membranes as they have ordering and condensing effects on lipids17, 18 and regulate fluidity and the phase behavior of membranes. 19, 20 Therefore, to be realistic, the models of plasma membranes should include cholesterol molecules in high concentration. This is also crucial when considering the effect of microplastics on model cell membranes: cholesterol could affect the permeation and dissolution of polymer nanoparticles in the membrane and – what is more important – nanosized microplastics could perturb the cholesterol distribution. To the best of our knowledge, such atomistic computational studies are still lacking.
In this paper, we use atomic-scale molecular dynamics simulations to evaluate the impact of nanosized microplastic particles on the properties of model cell membranes containing cholesterol molecules. To this end, we focused on a binary mixture of phospholipids and cholesterol. The primary goal is to understand how polymer nanoparticles affect the organization of cholesterol molecules within the lipid membrane. Our results clearly show that microplastics affect both the lateral and transverse distributions of cholesterol, thereby drastically disturbing the structure of the model cell membranes, which could have severe consequences for living cells.
COMPUTATIONAL METHODS
We performed atomic-scale molecular dynamics simulations of polypropylene and polystyrene nanoparticles (~ 4. 5 – 5. 0 nm in diameter) interacting with model cell membranes. We considered palmitoyl-oleoyl-phosphatidylcholine (POPC)-cholesterol and pure POPC membranes. All membrane systems consisted of 256 lipids; the POPC: cholesterol ratio was set to 2: 1 (33 mol % of cholesterol). The CHARMM36 force field21 and the CHARMM Generalized Force Field (CGenFF)22 were used for lipids and polymers, respectively. Simulations were performed in the NPT ensemble (at a temperature of 310 K and pressure of 1 bar) using the Gromacs 2023 suite. 23 The v-rescale thermostat24 and the C-rescale barostat25 were employed. The Lennard-Jones interactions were cut off at 1. 2 nm; the forces were smoothly switched to zero between 1. 0 and 1. 2 nm. The particle- mesh Ewald (PME) method was used to handle electrostatic interactions. 26 The time step was set to 2 fs. The initial structures of polymer chains were generated using the CHARMM-GUI Polymer Builder, 27, 28 while the CHARMM-GUI Membrane Builder29 was used to generate POPC-cholesterol and POPC lipid membranes. To evaluate the free energy of partitioning of a polymer nanoparticle from water into the lipid membrane, the umbrella sampling technique was used;30 a distance between the centers of mass of a nanoparticle and a membrane was chosen as a reaction coordinate. 31 The pull code of the Gromacs suite23 was used to generate starting configurations for umbrella sampling. After initial pulling, 40 windows were extracted with the reaction coordinate in the range of 1. 0 – 5. 0 nm; the spacing between windows was 0. 1 nm. Each window was simulated for 100 ns with the force constant set to 3000 kJ mol−1 nm−2; the last 80 ns were used for free-energy calculations. The Gromacs implementation23, 32 of the weighted histogram analysis (WHAM) method33 was employed to evaluate the free energy (potential of mean force). Statistical errors were estimated using bootstrapping analysis. 32 Simulation analysis was performed using in-house codes, the MEMBPLUGIN for VMD, 34 and standard Gromacs routines. 23 All snapshots were made using the VMD visualization software. 35 Further details of computational methods can be found in the Supporting Information.

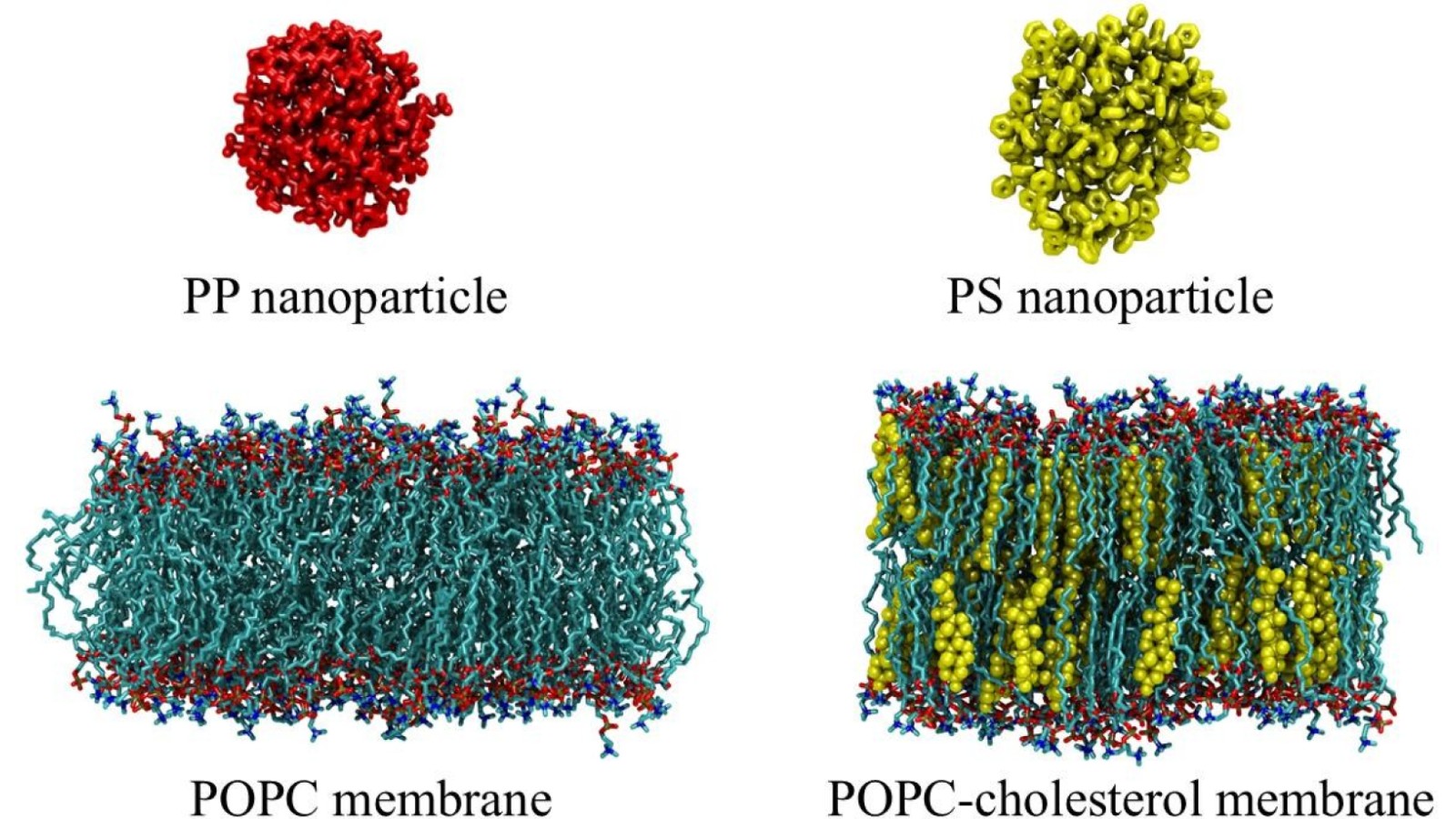
Figure 1. Components of the microplastics-membrane systems considered in the study. Shown are polypropylene and polystyrene nanoparticles as well as POPC- cholesterol and POPC lipid membranes. A detailed description of the microplastics- membrane systems can be found in the Supporting Information.
RESULTS AND DISCUSSION
For the purposes of our study, we considered lipid membranes consisting of a 2: 1 mixture of palmitoyl-oleoyl-phosphatidylcholine (POPC) and cholesterol (the cholesterol concentration was ~ 33 mol %) as well as pure POPC membranes, see Figure 1. As for nanosized microplastic particles, we focused on polypropylene (PP) and polystyrene (PS) nanoparticles, as both PP and PS are widely used in industry and households. The average size of the PP and PS particles was around 4. 0 – 4. 5 nm, i. e., the size of microplastics was comparable with the thickness of the lipid membrane. The full description of the systems is presented in the Supporting Information (Section S1, Tables S1 and S2).
First, we performed unbiased simulations of POPC-cholesterol and POPC membranes with a polymer nanoparticle (PP or PS) placed in the water phase. It turned out that nanosized microplastics do not adsorb on the membrane surface on a microsecond timescale, see Figure S1. Over the course of simulations, polymer nanoparticles made several attempts to bind to the membrane surface but were eventually pushed back into bulk water.
To evaluate the time required for a polymer nanoparticle to enter the lipid membrane, we calculated the free energy for partitioning of a polymer nanoparticle from the water phase to the center of the lipid membrane, see Figure 2. The free energy curves clearly show the existence of an energy barrier for each membrane system at hand. The nature of this barrier is in the unfavorable interactions between hydrophobic microplastics and hydrophilic head groups of phosphatidylcholine lipids or/and hydroxyl groups of cholesterol. For pure POPC bilayers, the height of such a barrier ΔG (measured with respect to bulk water) amounts to 67. 4±11. 4 and 33. 9±8. 9 kJ/mol for PP and PS microplastics, respectively, see Table 1; the corresponding peaks are located at 2. 66 and 2. 71 nm. Interestingly, the energy barrier for a PS nanoparticle is 2 times lower than that for a PP counterpart, despite the fact that the PS particle is a bit larger, see Table S2. Furthermore, the free energy profile of a PS nanoparticle within lipid/water interface is wider compared to a PP particle and includes more local peaks and minima, see Figure 2. The latter could be a sign of structural rearrangements of large phosphatidylcholine head groups upon their interactions with bulky phenyl side groups of PS chains. Replacing one third of POPC lipids with cholesterol molecules increases the free energy barrier for permeation of a PS nanoparticle to 53. 1±2. 4 kJ/mol (the peak is located at 3. 43 nm), see Table 1, most likely due to unfavorable interaction of PS chains with cholesterol. This effect is not observed for PP microplastics. The free energy barrier for partitioning a PP nanoparticle into the POPC-cholesterol amounts to 61. 3±0. 4 kJ/mol (the maximum is located at 3. 60 nm), i. e., changes slightly compared to a pure POPC membrane, given the large fluctuations of the free energy for a cholesterol-free system, see Figure 2(A).

Figure 2. Free energy profile for partitioning polypropylene (A) and polystyrene (B) nanoparticles from the water phase to the center of POPC-cholesterol and POPC membranes as a function of the distance between the centers of mass of a nanoparticle and a membrane. Statistical errors were estimated with the use of bootstrapping analysis.
Table 1. Thermodynamic, structural, and dynamic characteristics for considered membrane systems: POPC membranes with PP and PS nanoparticles (the POPC-PP and POPC-PS systems), POPC-cholesterol membranes with PP and PS nanoparticles (the POPC-CHOL-PP and POPC-CHOL-PS systems), and polymer-free POPC POPC-cholesterol membranes (the POPC and POPC-CHOL systems). Shown are free energy barriers ΔG for partitioning PP and PS nanoparticles into POPC and POPC-cholesterol membranes; errors were estimated using bootstrapping analysis. For lipid membranes with pre-inserted nanoparticles, presented are the area per lipid, the membrane thickness, the orientation of POPC head groups φPN and cholesterol molecules φCHOL with respect to the membrane normal, and the lateral diffusion coefficients for POPC lipids (DPOPC) and cholesterol (DCHOL). The results for polymer-free POPC and POPC-cholesterol membranes are shown as a reference. The average values were calculated using the last microsecond of 10 µs trajectories; errors are the standard deviations calculated over 10 ns blocks.

Using the free energy barriers ΔG for the nanoparticle partitioning, we can make a rough estimate for the rate of the passive nanoparticle permeation into the lipid membrane. Our estimates, based on the Kramers theory (see Section S1 in the Supporting Information), show that for the POPC-PS system with the lowest free energy barrier, the permeation rate is ~4·10 s-1, so that the nanoparticle permeation takes around 25 milliseconds. In turn, for the POPC-PP system with the highest barrier, one has ~10-4 s-1 for the permeation rate, i. e., a polymer nanoparticle needs around 3 hours to permeate. Obviously, such time scales are not accessible with atomic-scale computer simulations. It is important to point out that earlier coarse- grained simulations of PS nanoparticles interacting with POPC lipid membranes reported the permeation of a polymer nanoparticle into the phospholipid membrane on a time scale of a few microseconds. 7 Although the free energy of transfer of PS monomers and trimers from water to the POPC membrane was validated using atomistic models, 7 a coarse-grained PS nanoparticle as a whole partitions into the lipid membrane much faster than in atomic-scale simulations, most likely due to smoother interactions inherent to coarse-grained models. (36)
Figure 3. Radius of gyration of polypropylene (A) and polystyrene (B) nanoparticles in POPC and POPC-cholesterol membranes as a function of time. Top view of membrane systems with a PP nanoparticle inserted into POPC (C) and POPC- cholesterol (D) membranes and with a PS nanoparticle inserted into POPC (E) and POPC-cholesterol (F) membranes. For each system shown are the snapshots in the beginning and at the end of 10 µs simulations. Lipid membranes are shown in grey; PP and PS nanoparticles are shown in red and yellow, respectively.
Since atomic-scale simulations do not allow us to follow the process of partitioning microplastics into the lipid membrane, we chose to explore the behavior of a polymer nanoparticle within the membrane’s hydrophobic core after the partition step is completed. To this end, we considered a lipid membrane with a pre- inserted plastic particle and performed a 10-µs-long unbiased simulation run. In Figure 3(A, B), we present the radius of gyration Rg of a polymer nanoparticle in the interior of the membrane. For polypropylene nanoparticles, Rg increases over the course of simulations for both POPC-cholesterol and POPC membranes. Such a behavior indicates the dissolution of a nanoparticle within the membrane. This can also be confirmed via visual inspection of the snapshots taken in the beginning and at the end of 10 µs simulations, see Figure 3(C, D) for the top view and Figure S2(A, B) for the side view of microplastics-membrane systems. Importantly, cholesterol molecules considerably slow down the disintegration of a polypropylene nanoparticle: the radius of gyration of a PP particle in the POPC-cholesterol membrane is twice smaller than that in the cholesterol-free membrane after 10 µs of simulations. This finding is in line with the well-known ability of cholesterol molecules to slow down the lateral mobility in the membrane. 37 Polystyrene nanoparticles turned out to be more robust within the POPC membrane compared to PP microplastics: a PS particle started to dissolve in a cholesterol-free membrane only after 4 µs, see Figures 3(B, E) and S2(C). The nature of this robustness could be in the difference in interactions of polypropylene and polystyrene with hydrocarbon lipid tails. Analysis of atom-atom radial distribution functions of POPC acyl chains and polymer chains (see Figure S3) clearly shows that for polystyrene such interactions are less favorable, probably due to aromatic styrene side groups. In turn, cholesterol slows down the dissolution of a PS nanoparticle even further: adding 33 mol % of cholesterol to a phosphatidylcholine membrane suppresses the PS microplastics disintegration over the course of 10 µs simulations, see Figures 3(B, F) and S2(D). We note that none of the considered polymer nanoparticles dissolved completely within the membrane environment in atomic-scale simulations on a time scale of 10 microseconds. Individual polymer chains, on the other hand, turned out to be well equilibrated upon deformation and dissolution of nanoparticles, as seen from their radii of gyration and end-to-end distances (Figure S4). To assess the impact of microplastics on model cell membranes, we calculated a wide range of the membrane’s structural and dynamic characteristics using the last microsecond of 10 µs trajectories. As seen from Table 1, the insertion of a polymer nanoparticle into the membrane does not affect the average area per lipid as well as the orientation of POPC head groups and cholesterol molecules with respect to the membrane normal. However, microplastics increase the membrane thickness (measured as a distance between phosphate groups on the opposite leaflets), probably due to the presence of polymer chains in the middle of the membrane. The relative increase is around 0. 25 nm and turns out to be the same for both POPC- cholesterol and POPC systems, see Table 1. It is also instructive to stress that the average diameter of polymer nanoparticles considered in our study (see Table S2) is very close to the membrane thickness.
A more detailed insight into the microplastics-induced structural changes can be gained from component-wise mass density profiles along the bilayer normal, see Figure S5. A polymer nanoparticle is localized in the central part of a membrane. In fact, acyl chains of POPC lipids are substituted by the polymer in the middle of a membrane: upon microplastics insertion, the density of phosphatidylcholine lipids becomes systematically lower in the membrane center, see Figure S5. However, the most striking feature here is observed for cholesterol: although the density profiles for cholesterol molecules and polymer chains overlap in the center of a membrane, the density profiles for cholesterol molecules remain almost unchanged compared to the microplastics-free system, see Figure S5(D-F). The only explanation for such unusual behavior is in the lateral reorganization of cholesterol within the membrane: cholesterol molecules avoid unfavorable contacts with polymer chains and diffuse to the membrane regions with no polymer chains beneath. To prove that, we calculated component-wise mass density profiles along the direction perpendicular to the bilayer normal, 38 see Figure 4(A, B). Indeed, the maximum in the microplastic density can directly be linked to the minimum of cholesterol density, indicating the cholesterol depletion in the membrane domains above the polymer. This conclusion is also supported by visual inspection of the system snapshots, see Figure 4(C, D).

Figure 4. Component-wise mass density profiles for POPC-cholesterol membranes with polypropylene (A) and polystyrene (B) nanoparticles along the direction perpendicular to the membrane normal. The profiles were centered relative to the center of mass of a nanoparticle; the distance is measured from the edge of a membrane. Top view of POPC-cholesterol membranes with pre-inserted polypropylene (C) and polystyrene (D) nanoparticles at the end of 10 µs simulations. POPC lipids are shown in grey; PP and PS nanoparticles are shown in red and yellow, respectively. Cholesterol molecules in the upper leaflet are shown in green; the rest of the cholesterol is not shown for clarity.
To further characterize the microplastics-induced lateral reorganization of cholesterol molecules, we analyzed several radial distribution functions (RDFs) for cholesterol. As seen in Figure S6(A), the peak of the RDF for cholesterol-POPC becomes higher in the presence of a polymer nanoparticle, which implies stronger cholesterol-phosphatidylcholine interactions. As mentioned above, microplastics induce the formation of cholesterol-depleted domains, which should lead to a local increase in cholesterol concentration outside these domains and, correspondingly, to stronger cholesterol-POPC interactions. The same also applies to cholesterol- cholesterol interactions when polymer chains are present in the middle of a membrane, see Figure S6(B). As for the cholesterol-polymer RDF shown in Figure S6(C), it does not develop any noticeable peak, which indicates a lack of cholesterol- microplastics interactions. It is also instructive to examine the deuterium order parameter of POPC acyl chains. In cholesterol-free systems, microplastics do not affect the ordering of acyl chains as flexible POPC tails are able to easily adapt to polymer chains, see Figure S7(A, B). However, for the membrane systems with cholesterol, the situation becomes more involved. Cholesterol molecules themselves induce phospholipid tail ordering, 37 as it is easily seen in Figure S7. The insertion of polymer nanoparticles in POPC-cholesterol membranes results in a certain disordering of POPC acyl chains, which can be explained by the appearance of cholesterol-free domains in the membrane. Finally, inserting polymer nanoparticles into the interior of a lipid membrane leads to a systematic drop in the lateral diffusion of both phospholipids and cholesterol molecules, see Table 1. This effect is in line with earlier results7, 8 and could be associated with polymer chains entangled with phosphatidylcholine lipid tails and hinder lipid mobility. Interestingly, the nanoparticle’s impact on the lateral diffusion of lipids is stronger in the case of polystyrene: PS chains have bulky side groups and are characterized by slower conformational changes within the membrane compared to PP. In addition, one could expect an effect of a local increase in cholesterol concentration, which is known to slow down the lipid lateral mobility. 37
Overall, our findings clearly show that microplastics induce the lateral reorganization of cholesterol molecules in the membrane. The driving force of such reorganization is in unfavorable interactions of cholesterol molecules with polymer chains, but what makes the cholesterol-polymer interactions unfavorable? Our hypothesis is that the stiffness of a cholesterol molecule is responsible for such behavior. While flexible acyl chains of phospholipids are able to accommodate polymer chains in the middle of the membrane, cholesterol is too rigid to do the same. As a result, the polymer pushes cholesterol molecules towards the water phase, which is energetically unfavorable. To avoid this, cholesterol molecules laterally diffuse away from the polymer.
Although a phosphatidylcholine-cholesterol bilayer considered in our study is a very simplified model of cell membranes, the reported impact of microplastics could be of generic nature. Real membranes consist of many types of lipids that differ in the chain unsaturation and in the head group composition and charge. 39 However, as mentioned in the Introduction, a common feature of eukaryotic cell membranes is a high concentration of cholesterol molecules. Our findings suggest that lateral reorganization of cholesterol should not be affected to a significant extent by a particular lipid environment, as such reorganization is due to cholesterol- polymer interactions. Indeed, similar polystyrene-induced lateral redistribution of cholesterol was observed earlier in coarse-grained computer simulations of multicomponent lipid membranes. 7
Besides the lateral reorganization of cholesterol molecules, microplastics also perturb the cholesterol transverse distribution: in our atomic-scale simulations of microplastics-membrane systems, we observed several spontaneous cholesterol trans-bilayer motions (flip-flops), see Figure 5. Visual inspection shows that cholesterol flip-flops occur in the vicinity of polymer chains (or close to the edge of a polymer nanoparticle). Given that cholesterol molecules avoid contact with the polymer, the flip-flops can probably be linked to the structural defects induced by the movements of polymer chains in the interior of the membrane.

Figure 5. Time evolution of the positions of hydroxyl groups of flip-flopped cholesterol molecules in POPC-cholesterol membranes with polypropylene (A) and polystyrene (B) nanoparticles. The position is measured as a distance from the center of a membrane (dashed lines) along the membrane normal.
All in all, we witnessed six microplastic-induced flip-flop events in two POPC-cholesterol systems over 20 µs of simulations. This gives us the flip-flop rate of ~3·105 s-1. As cholesterol flip-flops are rarely observed in atomic-scale computer simulations, the flip-flop rate is often evaluated by calculating the free energy of transfer of a cholesterol molecule from a leaflet to the center of a membrane. 40 In particular, such calculations were performed for a bilayer with 98 POPC lipids and 2 cholesterol molecules, 41 with an estimate for the cholesterol flip-flop rate in the range 4. 8·102 – 4. 0·103 s-1. However, in our case, the cholesterol concentration in a POPC bilayer is much higher than that considered in ref 41 (33 versus 2 mol %). It is known that the cholesterol flip-flop rate decreases with cholesterol concentration. For instance, increasing cholesterol concentration in a dipalmitoyl- phosphatidylcholine bilayer from 3 to 20 mol % resulted in a drop of the flip-flop 15 rate by two orders of magnitude. 40 Therefore, we could consider a range of ~ 1 – 10 s-1 as a rough estimate for the cholesterol flip-flop rate in the microplastics-free POPC-cholesterol systems considered here. Comparing this value with ~3·105 s-1 measured in our atomistic simulations, we can conclude that microplastics speed up trans-bilayer motions of cholesterol molecules by ~ 4-5 orders of magnitude.
The cholesterol flip-flops are essential for the proper functioning of cell membranes, as flip-flops maintain the asymmetric distribution of lipids42 and balance the stresses in the opposite leaflets. 43 Furthermore, cholesterol molecules are also distributed asymmetrically in cytosolic and extracellular leaflets. 44, 45 Nanosized microplastics can destroy this asymmetry by transverse redistribution of cholesterol between leaflets; in turn, this could alter the lateral lipid organization in both leaflets. Therefore, the microplastics-induced speed-up of cholesterol flip-flops could have severe consequences for living cells.
CONCLUSIONS
In summary, here we presented the first atomic-scale computational study of the impact of nanosized microplastics on model cell membranes containing cholesterol, an essential component of membranes of eukaryotic cells. To this end, we focused on the interactions of polypropylene and polystyrene nanoparticles (~ 4. 0 – 4. 5 nm in diameter) with phosphatidylcholine-cholesterol lipid membranes. Our findings clearly show that, once a polymer nanoparticle partitions into the membrane interior, it alters both the lateral and transverse distributions of cholesterol within the membrane. In particular, microplastics induce the lateral reorganization of cholesterol molecules: because of its stiffness, cholesterol tends to avoid unfavorable contacts with polymer chains and diffuses away from the polymer. This results in the formation of domains depleted in cholesterol and has an immediate effect on the lipid packing and the phase behavior of a membrane. What is more,16 microplastics promote spontaneous trans-bilayer motions of cholesterol (flip-flops), most likely due to the structural defects induced by polymer chains in the membrane interior. Rough estimates indicate that polymer nanoparticles speed up cholesterol flip-flop by four-five orders of magnitude compared to polymer-free membrane systems. Overall, our computational findings suggest that nanosized microplastics could have a great impact on the distribution of cholesterol molecules in cell membranes and consequently alter the fluidity and the phase behavior of membranes, the transmembrane cholesterol distribution, and the ability of membranes to balance the stress.
ACKNOWLEDGMENTS
This work was supported by the Ministry of Science and Higher Education of the Russian Federation (state contract no. 075-15-2024-629, MegaGrant). All simulations were performed using computing facilities of Novgorod State University, Russia.
SUPPORTING INFORMATION
Detailed description of the models and methods, additional results for binding of microplastics to the lipid membrane, and for the membranes with pre-inserted polymer nanoplastics.
REFERENCES
Thompson, R. C. ; Olsen, Y. ; Mitchell, R. P. ; Davis, A. ; Rowland, S. J. ; John, A.W. G. ; McGonigle, D. ; Russell, A. E. Lost at Sea: Where is all the plastic? Science 2004, 304, 838–838. DOI: 10. 1126/science. 1094559.
Thompson, R. C. ; Courtene-Jones, W. ; Boucher, J. ; Pahl, S. ; Raubenheimer, K. ; Koelmans. A. A. Twenty years of microplastic pollution research – what have we learned? Science 2024, 386, eadl2746. DOI: 10. 1126/science. adl2746
Banerjee, A. ; Shelver, W. L. Micro- and nanoplastic induced cellular toxicity in mammals: a review. Sci. Total Environ. 2021, 755, 142518. DOI: 10. 1016/j. scitotenv. 2020. 142518
Liu, Z. ; You, X. Recent progress of microplastic toxicity on human exposure basedon in vitro and in vivo studies. Sci. Total Environ. 2023, 903, 166766. DOI: 10. 1016/j. scitotenv. 2023. 166766
Gigault, J. ; El Hadri, H. ; Nguyen, B. ; Grassl, B. ; Rowenczyk, L. ; Tufenkji, N. ; Feng, S. ; Wiesner, M. Nanoplastics are neither microplastics nor engineered nanoparticles. Nat. Nanotechnol. 2021, 16, 501-507. DOI: 10. 1038/s41565-021-00886-4
Zhao, J. ; Stenzel, M. H. Entry of nanoparticles into cells: the importance of nanoparticle properties. Polym. Chem. 2018, 9, 259-272. DOI: 10. 1039/C7PY01603D
Rossi, G. ; Barnoud, J. ; Monticelli, L. Polystyrene nanoparticles perturb lipid membranes. J. Phys. Chem. Lett. 2014, 5, 241-246. DOI: 10. 1021/jz402234c
Bochicchio, D. ; Panizon, E. ; Monticelli, L. ; Rossi, G. Interaction of hydrophobic polymers with model lipid bilayers. Sci. Rep. 2017, 7, 6357. DOI: 10. 1038/s41598- 07-06668-0
Li, L. ; Li, S. ; Xu, Y. ; Ren, L. ; Yang, L. ; Liu, X. ; Dai, Y. ; Zhao, J. ; Yue, T.Distinguishing the nanoplastic–cell membrane interface by polymer type and aging properties: translocation, transformation and perturbation. Environ. Sci.: Nano 2023, 10, 440-453. DOI: 10. 1039/D2EN00800A
Hollóczki, O. ; Gehrke, S. Can nanoplastics alter cell membranes? ChemPhysChem 2020, 21, 9-12. DOI: 10. 1002/cphc. 201900481
Bochicchio, D.; Cantu, L. ; Cadario, M. V. ; Palchetti, L. ; Natali, F. ; Monticelli, L. ; Rossi, G. ; Del Favero, E. Polystyrene perturbs the structure, dynamics, and mechanical properties of DPPC membranes: An experimental and computational study. J. Colloid Interf. Sci. 2022, 605, 110-119. DOI: 10. 1016/j. jcis. 2021. 07. 069
Wang, W.; Zhang, J. ; Qiu, Z. ; Cui, Z. ; Li, N. ; Li, X. ; Wang, Y. ; Zhang, H. ; Zhao, C. Effects of polyethylene microplastics on cell membranes: A combined study of 18 experiments and molecular dynamics simulations. J. Hazard. Mater. 2022, 429, 128323. DOI: 10. 1016/j. jhazmat. 2022. 128323
Yuan, S. ; Zhang, H. ; Yuan, S. Understanding the transformations of nanoplastic onto phospholipid bilayers: Mechanism, microscopic interaction and cytotoxicity assessment. Sci. Total Environ. 2023, 859, 160388. DOI: 10. 1016/j. scitotenv. 2022. 160388
Cheng, S. ; Ye, Z. ; Wang, X. ; Lian, C. ; Shang, Y. ; Liu, H. The effects of adsorbed benzo(a)pyrene on dynamic behavior of polystyrene nanoplastics through phospholipid membrane: A molecular simulation study. Colloids Surf. B:Biointerfaces 2023, 224, 113211. DOI: 10. 1016/j. colsurfb. 2023. 113211
Van Meer G. ; Voelker D. R. ; Feigenson G. V. Membrane lipids: where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112-24. DOI: 10. 1038/nrm2330
Sackmann, E. Biological membranes architecture and function, in: R. Lipowsky, E. Sackmann (Eds. ), Structure and Dynamics of Membranes, Elsevier, Amsterdam, 1995, pp. 1-64.Hung, W. -C. ; Lee, M. -T. ; Chen, F. -Y. ; Huang, H. W. The condensing effect of cholesterol in lipid bilayers. Biophys. J. 2007, 92, 3960-3967. DOI: 10. 1529/biophysj. 106. 099234
Urbina, J. A. ; Pekerar, S. ; Le, H. ; Patterson, J. ; Montez, B. ; Oldfield, E. Molecular order and dynamics of phosphatidylcholine membranes in the presence of cholesterol, ergosterol and lanosterol: a comparative study using 2H, 13C and 31P NMR spectroscopy. Biochim. Biophys. Acta 1995, 1238, 163-176. DOI: 10. 1016/0005-2736(95)00117-L
Ipsen, J. H. ; Karlström, G. ; Mourtisen, O. G. ; Wennerström, H. ; Zuckermann, M. J. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta 1987, 905, 162-172. DOI: 10. 1016/0005-2736(87)90020-4
Sezgin, E. ; Levental, I. ; Mayor, S. ; Eggeling, C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361-374. DOI: 10. 1038/nrm. 2017. 16
Klauda, J. B. ; Venable, R. M. ; Freites, J. A. ; O’Connor, J. W. ; Tobias, D. J. ; Mondragon-Ramirez, C. ; Vorobyov, I. ; MacKerell, A. D., Pastor, R. W. Update of the CHARMM all-atom additive force field for lipids: validation on six lipid types. J. Phys. Chem. B 2010, 114, 7830-7843. DOI: 10. 1021/jp101759q
Vanommeslaeghe, K. ; Hatcher, E. ; Acharya, C. ; Kundu, S. ; Zhong, S.; Shim, J. ; Darian, E. ; Guvench, O. ; Lopes, P. ; Vorobyov, I. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2010, 31, 671-690. DOI: 10. 1002/jcc. 21367 19 Páll, S. ; Zhmurov, A. ; Bauer, P. ; Abraham, M. ; Lundborg, M. ; Gray, A. ; Hess, B. ; Lindahl, E. Heterogeneous parallelization and acceleration of molecular dynamics simulations in GROMACS. J. Chem. Phys. 2020, 153, 134110. DOI: 10. 1063/5. 0018516
Bussi, G. ; Donadio, D. ; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. DOI: 10. 1063/1. 2408420
Bernetti, M. ; Bussi, G. Pressure control using stochastic cell rescaling. J. Chem.Phys. 2020, 153, 114107. DOI: 10. 1063/5. 0020514
Essman, U. ; Perera, L. ; Berkowitz, M. L. ; Darden, T. ; Lee, H. ; Pedersen, L. G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577-8592. DOI: 10. 1063/1. 470117 Jo, S. ; Kim, T. ; Iyer, V. G. ; Im, W. CHARMM-GUI: a web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859-1865. DOI: 10. 1002/jcc. 20945
Choi, Y. K. ; Park, S. J. ; Park, S. ; Kim, S. ; Kern, N. R. ; Lee, J. ; Im, W.CHARMM-GUI polymer builder for modeling and simulation of synthetic polymers. J. Chem. Theory Comput. 2021, 17, 2431-2443. DOI: 10. 1021/acs. jctc. 1c00169
Wu, E. L. ; Cheng, X. ; Jo, S. ; Rui, H. ; Song, K. C. ; Davila-Contreras, E. M. ; Qi,Y. F. ; Lee, J. M. ; Monje-Galvan, V. ; Venable, R. M. ; Klauda, J. B. ; Im, W. CHARMM-GUI Membrane Builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997-2004. DOI: 10. 1002/jcc. 23702
Torrie, G. M. ; Valleau, J. P. Nonphysical sampling distributions in Monte Carlo free-energy estimation: Umbrella sampling. J. Comput. Phys. 1977, 23, 187-199.DOI: 10. 1016/0021-9991(77)90121-8
Gurtovenko, A. A. Molecular-level insight into the interactions of DNA- polycation complexes with model cell membranes. J. Phys. Chem. B 2019, 123, 6505-6514. DOI: 10. 1021/acs. jpcb. 9b05110
Hub, J. S. ; de Groot, B. L. ; van der Spoel, D. g_wham - a free weighted histogram analysis implementation including robust error and autocorrelation estimates. J. Chem. Theory Comput. 2010, 6, 3713-3720. DOI: 10. 1021/ct100494z
Kumar, S. ; Rosenberg, J. M. ; Bouzida, D. ; Swendsen, R. H. ; Kollman, P. A. The weighted histogram analysis method for free energy calculations on biomolecules. I. The method. J. Comput. Chem. 1992, 13, 1011-1021. DOI: 10. 1002/jcc. 540130812
Guixà-González, R. ; Rodríguez-Espigares, I. ; Ramírez-Anguita, J. M. ; Carrió- Gaspar, P. ; Martinez-Seara, H. ; Giorgino, T. ; Selent, J. MEMBPLUGIN: studying membrane complexity in VMD. Bioinformatics 2014, 30, 1478-1480. DOI: 10. 1093/bioinformatics/btu037 20 Humphrey, W. ; Dalke, A. ; Schulten, K. VMD - Visual Molecular Dynamics. J.Mo lec. Graphics 1996, 14, 33-38. DOI: 10. 1016/0263-7855(96)00018-5
Marrink, S. J. ; Risselada, H. J. ; Yefimov, S. ; Tieleman, D. P. ; de Vries, A. H. The Martini force field: coarse grained model for biomolecular simulations. J. Phys. Chem. B 2007, 111, 7812-7824. DOI: 10. 1021/jp071097f
Róg, T. ; Pasenkiewicz-Gierula, M. ; Vattulainen, I. ; Karttunen, M. Ordering effects of cholesterol and its analogues. Biochim. Biophys. Acta – Biomembranes 2009, 1788, 97-121. DOI: 10. 1016/j. bbamem. 2008. 08. 022
Risselada, H. J. ; Marrink, S. J. The molecular face of lipid rafts in model membranes. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 17367-17372. DOI: 10. 1073/pnas. 0807527105 Gennis, R. B. Biomembranes: Molecular Structure and Function. Springer- Verlag: New York, 1989. Bennett, W. F. D. ; MacCallum, J. L. ; Hinner, M. J. ; Marrink, S. J. ; Tieleman, D.P. Molecular view of cholesterol flip-flop and chemical potential in different membrane environments. J. Am. Chem. Soc. 2009, 131, 12714-12720. DOI: 10. 1021/ja903529f
Jo, S. ; Rui, H. ; Lim, J. B. ; Klauda, J. B. ; Im, W. Cholesterol flip-flop: insights from free energy simulation studies. J. Phys. Chem. B 2010, 114, 13342-13348. DOI: 10. 1021/jp108166k
Steck, T. L. ; Lange, Y. Transverse distribution of plasma membrane bilayer cholesterol: picking sides. Traffic 2018, 19, 750-760. DOI: 10. 1111/tra. 12586
Bruckner, R. J. ; Mansy, S. S. ; Ricardo, A. ; Mahadevan, L. ; Szostak, J. W. Flip- flop-induced relaxation of bending energy: implications for membrane remodeling. Biophys. J. 2009, 97, 3113-3122. DOI: 10. 1016/j. bpj. 2009. 09. 025
Marquardt, D. ; Geier, B. ; Pabst, G. Asymmetric lipid membranes: towards more realistic model systems. Membranes 2015, 5, 180-196. DOI: 10. 3390/membranes5020180
Falkovich, S. G. ; Martinez-Seara, H. ; Nesterenko, A. M. ; Vattulainen, I. ; Gurtovenko, A. A. What can we learn about cholesterol’s transmembrane distribution based on cholesterol-induced changes in membrane dipole potential? J. Phys. Chem. Lett. 2016, 7, 4585-4590. DOI: 10. 1021/acs. jpclett. 6b02123











![Микропластик в воде и пище: [не]осведомленность](/upload/iblock/3a0/h99p373wv55ur266lw4qbbegvp6f8033.jpg)